David Baltimore, ricercatore statunitense e premio nobel per la medicina nel 1975, ha influenzato il progresso scientifico con importanti ricerche sulla trascrittasi inversa, sull’immunologia e sulla ricombinazione del DNA. Baltimore ha inoltre lavorato su una classificazione dei virus che, in base al genoma e alla modalità di replicazione, vengono collocati in 7 diverse classi.
David Baltimore
David Baltimore è un biologo ricercatore nato il 7 marzo 1938 a New York. È stato premio Nobel per la Medicina nel 1975 insieme a Howard M. Temin e Renato Dulbecco per le loro scoperte riguardo l’interazione tra i virus e il materiale genetico della cellula. Attualmente è presidente emerito al California Institute of Technology (Caltech), dove è stato presidente e professore di biologia dal 1997 al 2006. Recentemente Baltimore, insieme ad altri premi Nobel in ambito scientifico, è stato assunto dalla Altos Labs, azienda fondata da Jeff Bezos, al fine di lavorare ad un nuovo progetto il cui obiettivo è “riprogrammare” le cellule per invertire il processo di invecchiamento cellulare.

La trascrittasi inversa
Nel 1970 Baltimore, contemporaneamente a Temin, ha isolato e studiato la trascrittasi inversa, un enzima capace di trascrivere DNA a partire da un filamento stampo di RNA. In questo modo è riuscito a classificare una distinta categoria di virus, successivamente denominati retrovirus. I Retrovirus, come l’HIV, infettano la cellula ospite con il loro materiale genico costituito da un filamento di RNA ed utilizzano la trascrittasi inversa per sintetizzare dal loro RNA del DNA.
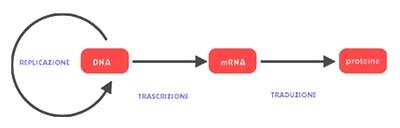
Questa scoperta ha così confutato il “dogma centrale” della biologia, che asseriva che l’informazione genica passasse solamente dal DNA all’RNA e successivamente da RNA a proteine.
La classificazione di Baltimore
Un virus è una particella infettiva intracellulare obbligata poiché capace di riprodursi solo all’interno di cellule viventi. Un virus è composto da materiale genetico a DNA o RNA (a singolo o a doppio filamento) e da un rivestimento proteico detto capside.
I virus non sono in grado di effettuare in autonomia nessun processo metabolico poiché non posseggono sistemi di biosintesi. Essi infatti contengono solo l’informazione genetica necessaria alla replicazione e alla predisposizione dei virioni. L’acido nucleico virale codifica enzimi necessari alla replicazione del materiale genico ed alla produzione di proteine strutturali che costituiscono il rivestimento del virus. Tutte le altre funzioni come produzione di energia e di complessi trascrizionali e traduzionali per la sintesi proteica, sono fornite dalla cellula infettata.
Nel tempo sono stati prodotti diversi sistemi di classificazione dei virus, ma nessuno è mai stato completamente esaustivo.
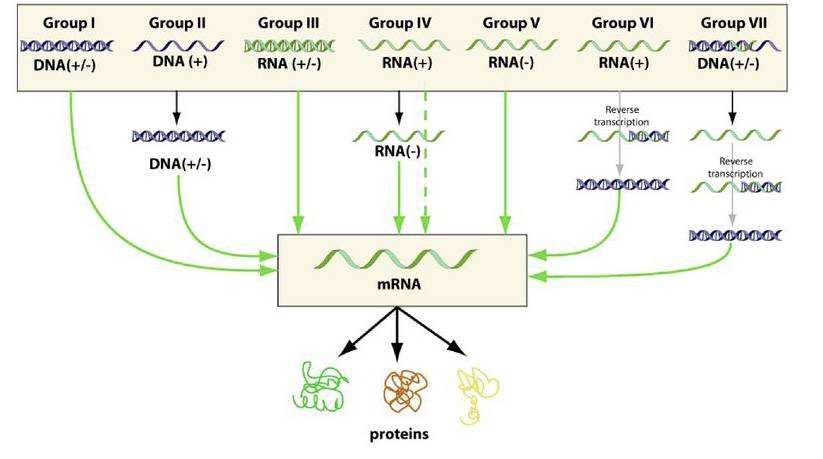
Baltimore ha messo a punto una classificazione che considera la natura dell’acido nucleico virale e il modo in cui da questo si ottengono le proteine virali. La classificazione consta di 7 classi di virus:
- DNA a doppia elica
- DNA a singolo filamento
- RNA a doppia elica
- Un filamento ad RNA con polarità positiva
- Un filamento con polarità negativa
- Singolo filamento di RNA con polarità positiva
- DNA a doppia elica presentante dei gap
Virus di classe I
Nella prima classe i virus presentano un genoma a doppia elica di DNA.
Fanno parte di questa classe Adenoviridae, Herpesviridae, Papillomaviridae, Polyomaviridae e Poxvioridae.
In particolare, gli Herpes virus aderiscono alla cellula ospite mediante un recettore specifico che permette loro di penetrare al suo interno. La loro penetrazione determina la fusione dell’envelope alla membrana immettendo al suo all’interno solo il nucleocapside virale. Le proteine del citoscheletro, veicolano il virus nel nucleo dove l’interazione con la membrana nucleare determina la completa decapsidazione del virione. Qui il virus si appropria degli apparati trascrizionali dell’ospite.
Dalla traduzione del genoma virale nel citoplasma si ottengono tutte le proteine virali, successivamente assemblate, producendo la progenie virale. L’Herpes virus per poter fuoriuscire dalla cellula ospite effettua un’esocitosi. Esocitando porta con sé una porzione di membrana plasmatica contenente le sue proteine di rivestimento che caratterizzano l’envelope.
Alcuni virioni possono migrare attraverso le terminazioni assoniche dell’ospite raggiungendo i gangli alla base del nervo trigemino e lì, perdendo tutti i rivestimenti, rimangono “extracromosomali” nel nucleo delle cellule nervose, producendo infezioni latenti dove non si replicano. Quando il sistema immunitario è debilitato, a causa di stress o nervosismo, il virus torna a proliferare sviluppando il famoso Herpes labiale.

Virus di classe II
I virus di seconda classe hanno un genoma a singola elica di DNA positiva o negativa. Usano la DNA polimerasi dell’ospite per trasformare il genoma in doppia elica di DNA. A questo punto possono trascrivere il loro materiale genico e tradurre le loro proteine grazie ai complessi appropriati forniti dall’ospite. Di questa classe fanno parte i Parvovirus, peculiari poichè da soli non sono in grado di espletare il proprio ciclo replicativo, ma devono necessariamente associarsi a dei virus Helper come Herpesvirus o Adenovirus. Pertanto, per poter intraprendere un’infezione, penetrano cellule già infettate da altri virus.

Virus di classe III
I virus di terza classe sono i Reovirus che presentano un genoma a doppia elica di RNA che si replica nel citoplasma dell’ospite.
I Reovirus, una volta che aderiscono alla membrana dell’ospite, effettuano un’endocitosi che produce degli endosomi al suo interno. Successivamente, questi endosomi nel citoplasma, si fondono con dei lisosomi che con il loro pH acido consentono la decapsidazione del virus.
Una volta che il genoma virale si trova all’interno del citoplasma, essendo una doppia elica a RNA, impiega una RNA polimerasi RNA-dipendente (portata appositamente con sé) per sfruttare l’elica antisenso come template per generare un mRNA da cui deriveranno tutte le sintesi proteiche. Una volta completato l’assemblaggio dei virioni il loro ciclo si completa con la lisi della cellula ospite.
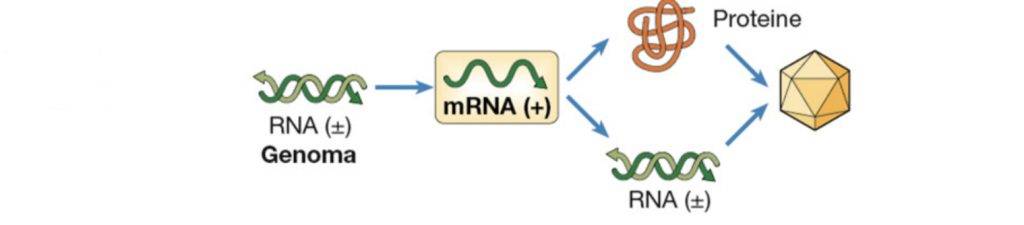
Virus di classe IV
Dei virus di quarta classe fanno parte i Coronavirus, Flavivirus e Picornavirus. Questi hanno un genoma a singola elica di RNA a polarità positiva. Si tratta di virus che, presentando un RNA direttamente traducibile, una volta entrati nel citoplasma riescono ad appropriarsi dell’apparato traduzionale dell’ospite impedendogli la traduzione dei propri messaggeri. Traducono per prima una RNA polimerasi RNA-dipendente in modo da poter replicare il loro genoma che può fungere da mRNA (per incrementare la sintesi proteica) oppure incapsidarsi nelle proteine di rivestimento generando un virus finito.
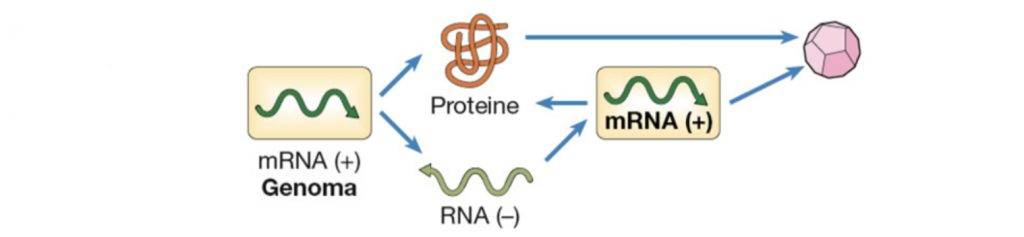
Virus di classe V
I virus di quinta classe sono caratterizzati da un RNA a singola elica a polarità negativa che si replica nel citoplasma. Di questa categoria fanno parte Orthomyxovirus, Rhabdovirus e Paramyxovirus.
Il Rhabdovirus è il virus della rabbia e lo si può contrarre attraverso la saliva di animali infetti.
Una volta che il suo materiale genetico entra nel citoplasma, essendo a RNA con polarità negativa, produce dei messaggeri con una RNA polimerasi RNA-dipendente. Una parte dei messaggeri a polarità positiva viene utilizzata per la sintesi proteica, mentre un’altra parte fa da filamento stampo per produrre nuovamente un filamento ad RNA negativo, successivamente incapsidato. Una volta che il nucleocapside si assembla, si dirige verso la regione della membrana dove sono presenti le sue glicoproteine di superficie e quindi, gemmando fuori dalla cellula, acquisisce l’envelope.
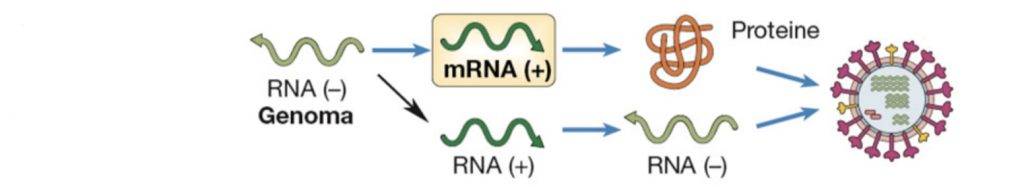
Virus di classe VI
La sesta classe è costituita dai Retrovirus. I Retrovirus sono virus a RNA con polarità positiva, ed una volta dentro l’ospite non sono direttamente traducibili.
Una volta che un Retrovirus prende contatti con la superficie dell’ospite, penetra all’interno e a seguito della sua decapsidazione il materiale genetico nel citoplasma, è target di una Trascrittasi inversa.
Si tratta di un enzima che porta con sé e che è in grado di polimerizzare un filamento di DNA sfruttando come filamento stampo l’RNA che possiede. L’enzima genera un ibrido RNA-DNA, che con un dominio in grado di degradare lo stesso RNA genera un singolo filamento di DNA.
Il singolo filamento di DNA viene poi utilizzato come stampo per generare il suo complementare, fino ad arrivare ad una doppia elica.
Quest’ultima, grazie alle integrasi, riesce ad integrarsi nel genoma dell’ospite, replicando passivamente insieme ad esso. Nel momento in cui raggiunge una determinata fase del suo ciclo vitale, inizia la seconda fase preposta alla produzione della progenie virale. Il virus a questo punto inizia a trascrivere il suo mRNA e, traducendolo, sintetizza le sue proteine. Questa meccanismo permette ad un particolare tipo di retrovirus, l’HIV, di riconoscere uno specifico recettore per i linfociti T. Infatti, quando comincia la sua fase virale, degrada i linfociti T dell’ospite determinandone una forte compromissione del sistema immunitario.
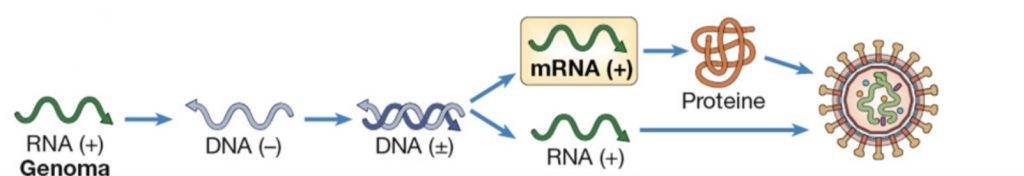
Virus di classe VII
L’ultima classe di virus presenta un genoma costituito da una molecola a doppio filamento di DNA che presenta dei gap in alcuni punti. Un esempio di tale virus è l’Hepadnavirus, che è in grado di generare epatiti.
Nel momento in cui l’Hepadnavirus aderisce all’ospite e penetra al suo interno, i sistemi di riparazione del DNA dello stesso, riempiranno i suoi gap e determineranno la circolarizzazione del genoma virale. Il virus raggiunge la sede nucleare ed utilizza la polimerasi dell’ospite per la propria replicazione. Successivamente sfrutta il complesso traduzionale nel citoplasma per effettuare la sintesi proteica. A questo punto il virus, quando deve produrre le copie di genoma virale da dover essere incapsidate, invece di sfruttare la DNA polimerasi dell’ospite, usa una sua trascrittasi inversa. Questa trascrittasi inversa sfrutta gli mRNA virali per generare un genoma a DNA. Dunque, mentre HIV nella seconda fase del suo ciclo passa da DNA a RNA, l’Hepadnavirus passa da RNA a DNA. Questa trascrittasi inversa determina quindi l’introduzione dei gap nel doppio filamento, tipici dei virus di classe 7.

Fonti immagini:
- https://it.wikipedia.org/wiki/David_Baltimore#/media/File:Dr._David_Baltimore2.jpg
- https://www.researchgate.net/figure/Figura-4-Struttura-della-trascrittasi-inversa-di-HIV-1_fig53_294736697
- http://www.cpsanalitica.com/biotecnologie/biologia_molecolare/biologia_molecolare.htm
- https://slideplayer.it/slide/13118959/
- https://documen.site/download/genomi-a-dna-herpesviridae-e_pdf
{kind=link}
Fonti bibliografiche:
- Dehò G., Galli E., Biologia dei Microrganismi, Casa Editrice Ambrosiana, Terza ed. 2018
- Fermin, G. (2018). Viruses: Molecular Biology, Host Interactions and Applications to Biotechnology. Elsevier.
- Lostroh, P. (2019). Molecular and Cellular Biology of Viruses. Garland Science
- https://www.bbe.caltech.edu/people/david-baltimore
- https://infinite.mit.edu/video/david-l-baltimore
- https://tech.everyeye.it/notizie/jeff-bezos-assume-migliori-scienziati-premio-nobel-sconfiggere-vecchiaia-565516.html