ll sistema del complemento è un elemento essenziale del sistema immunitario nei meccanismi di difesa umorale contro gli agenti infettivi; si tratta di un sistema costituito da diverse proteine circolanti nel sangue e di membrana, capaci di interagire reciprocamente e con le membrane cellulari. Viene definito “Complemento” per la capacità di potenziare (complementare) l’attività opsonizzante degli anticorpi.
- E’ un sistema di difesa innato ed inducibile.
- E’ attivato da una cascata enzimatica costituito da proteasi sieriche.
- Un ruolo di primaria importanza è rivestito dall’enzima C3 convertasi.
Il sistema del complemento collega l’immunità innata e quella acquisita tramite meccanismi come:
- Aumentando le risposte anticorpali e la memoria immunologica.
- Lisando le cellule estranee.
- Bonificando il microambiente da immunocomplessi e da cellule apoptotiche.
Scoperta e caratteristiche del sistema del complemento
Il termine complemento fu usato per la prima volta nel 1898 da Jules Bordet (Fig.1) che aveva notato che, incubando una miscela di batteri con siero contenente anticorpi rivolti contro i batteri stessi, questi venivano lisati (distrutti). Se invece il siero veniva riscaldato a una temperatura di 56 °C la lisi non avveniva. Poiché era stato già dimostrato che gli anticorpi resistevano al riscaldamento, Bordet ipotizzò la presenza nel siero di un componente termolabile che completava l’azione degli anticorpi e che battezzò appunto complemento.
Il sistema consiste di circa trenta proteine, sia solubili (plasmaproteine) che di membrana, presenti in tutti i liquidi organici, indicate con la lettera C seguita da numeri progressivi. In condizioni fisiologiche queste proteine sono inattive. Circa il 90% delle proteine del complemento vengono sintetizzate nel fegato mentre il restante 10% viene sintetizzato principalmente dai monociti, macrofagi e fibroblasti. Il complemento può essere attivato direttamente dai microbi, costituendo un sistema di difesa dell’immunità innata, oppure dagli anticorpi attaccati al microbo, costituendo un braccio effettore dell’immunità adattativa.

Attivazione del sistema del complemento
Come accade per altri sistemi operanti nell’organismo, come quello della coagulazione del sangue e quello delle chinine, l’attivazione del complemento avviene con un meccanismo detto “a cascata” per attivazione sequenziale dei vari componenti complementari che circolano in forma inattiva.
Esistono 3 vie per attivare il complemento, specificatamente sono:
- Classica
- Lectina
- Alternativa
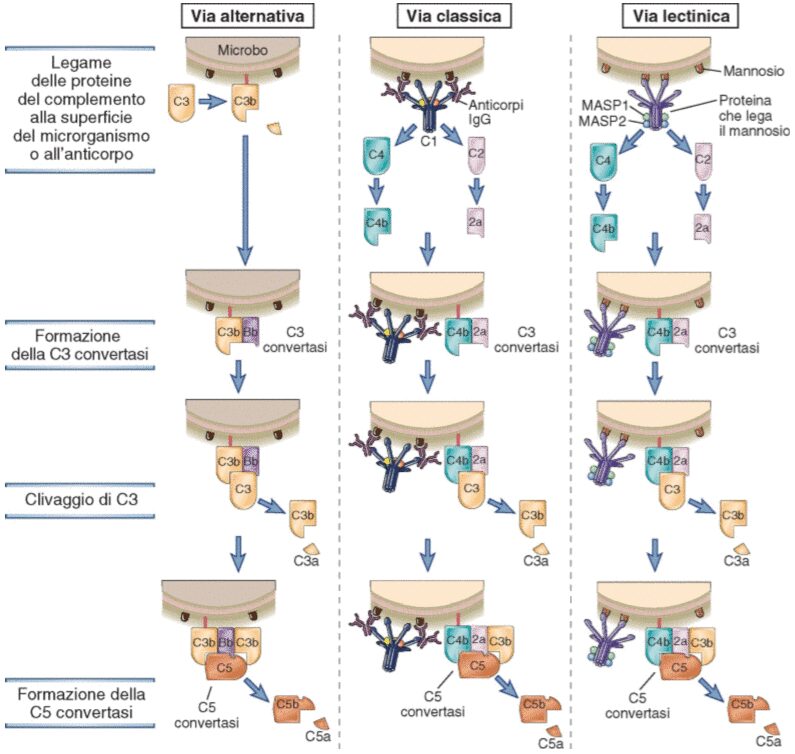
La via classica del complemento
La via classica è avviata dal legame della frazione C1 ai domini costanti delle immunoglobuline IgG e IgM complessate all’antigene. La frazione C1 è un è un complesso proteico composto dalle subunità C1q, C1r e C1s: la subunità C1q è deputata al legame con l’anticorpo mentre la C1r e C1s sono dotate di attività proteasica; la C1q deve legare specificatamente la regione Fc dell’anticorpo ed inoltre, per innescare il processo, è necessario che il legame avvenga solo con anticorpi legati all’antigene e non circolanti; poiché abbia inizio l’attivazione è necessario che due o più porzioni Fc siano accessibili alla frazione C1.
Il legame della subunità C1q alle regione Fc delle IgG o delle IgM porta all’attivazione della subunità C1r, che cliva la subunità C1s attivandola, a sua volta, la C1s attivata va ad agire sulla componente successiva della cascata del complemento, la frazione C4. Si genera così un frammento più piccolo C4a rilasciato nella fase fluida ed un frammento grande C4b che si legherà sulla superficie microbica. Un’altra componente importante è la frazione C2, che si lega al frammento C4b e, a questo punto, viene clivato da C1s generando un frammento C2b solubile ed il frammento C2a che rimane fisicamente associato al frammento C4b legato sulla superficie microbica. Il complesso C4b2a rappresenta la C3 convertasi della via classica, dotata della capacità di legare e clivare la frazione C3.
La via Lectina
La via lectinica di attivazione del sistema del complemento si innesca in modo anticorpo-indipendente a partire dall’interazione dei polisaccaridi microbici con le lectine circolanti come la lectina legante il mannosio (MBL).
Le lectine solubili fanno parte della famiglia delle collectine e da un punto di vista strutturale sono molto simili alla subunità C1q della frazione C1. La MBL si lega ai residui di mannosio dei polisaccaridi grazie a domini specifici presenti sull’estremità C-terminale che ha la capacità di riconoscere molecole di natura saccaridica. La MBL interagisce con le serine proteasi associate, le MASP; i cui membri sono: MASP-1, MASP-2 e MASP-3. Le proteine MASP hanno una struttura omologa a quella delle proteasi C1r e C1s della frazione C1 e svolgono funzioni molto simili. La proteina MASP-1 può formare un complesso tetramerico con MASP-2 simile a quello formato dalle subunità C1r e C1s; nello specifico MASP-2 rappresenta la proteasi che va a clivare le frazioni C4 e C2. Gli eventi conseguenti a questa reazione sono gli identici a quelli che avvengono nella via classica.
La via alternativa complemento
La via alternativa di attivazione del sistema del complemento porta alla proteolisi della frazione C3 e alla conseguente formazione del frammento C3b che si lega alla superficie microbica. Quando viene tagliato si formano due frammenti, il frammento C3b che espone il suo gruppo tioesterico reattivo ed il frammento C3a, più piccolo, che rientra in circolo. Il gruppo tioesterico del C3b ha la capacità di legarsi covalentemente sulla superficie dei microrganismi. Questo legame induce l’esposizione di un ulteriore sito di legame per un’altra proteina plasmatica denominata fattore B. Il fattore B si lega al frammento C3b e viene clivato a opera di una serina proteasi plasmatica chiamata fattore D.
Il clivaggio genera un piccolo frammento denominato Ba ed un frammento più grande chiamato Bb che rimane legato al frammento C3b. Il complesso C3bBb rappresenta la C3 convertasi della via alternativa e la sua funzione è clivare massicciamente la frazione C3 presente nel plasma, innescando così un circuito di amplificazione che genera quantità sempre maggiori di C3b. I frammenti C3b, quindi, possono legarsi alla superficie del microbo oppure legarsi alla C3 convertasi stessa portando alla formazione di un complesso contenente un frammento Bb e due frammenti C3b. Questo complesso si definisce come C5 convertasi.
Attivazione del complesso litico
Come illustrato (Fig.2), le tre vie di attivazione del sistema del complemento, la classica, l’alternativa e la lectinica, portano alla costituzione di due complessi diversi, il C4b2a e il C3bBb, ma con un’identica funzione: quella di scindere il C3 in due frammenti, C3a e C3b, e di legarsi al C3b costituendo le C5 convertasi.
Le C5 convertasi originate innescano l’attivazione delle componenti finali del sistema del complemento che conducono alla formazione del MAC (Membrane Attack Complex) e alla distruzione della cellula. Le C5 convertasi clivano il C5 in due frammenti: C5a che rimane in soluzione espletando importanti funzioni, e il C5b che si lega alle altre proteine del complemento già presenti sulla membrana, le C6 e C7 formando il complesso C5b,6,7. Una volta legata, C7 si inserisce nel doppio strato lipidico della membrana microbica dove diventa un recettore per C8 che a sua volta legandosi va a definire il complesso C5b,6,7,8.
L’ultima tappa per la formazione di un MAC attivo è il legame con C9 che in associazione al complesso C5b-8 polimerizza e forma un poro nella membrana batterica in modo simile alla perforina rilasciata dai linfociti T citotossici e NK. Attraverso i pori prodotti dalla polimerizzazione del C9, l’acqua e gli ioni hanno libero accesso all’interno della cellula. Ciò ne determina dapprima il rigonfiamento osmotico e poi lo scoppio della stessa.
Attività biologiche del sistema del complemento
Le molecole del sistema del complemento hanno altre funzioni immunitarie che sono mediate dai recettori del complemento localizzate su varie cellule.
- Il CR1 (CD35) promuove la fagocitosi e partecipa all’eliminazione degli immunocomplessi.
- Il CR2 (CD21) regola la produzione di anticorpi da parte delle cellule B.
- I recettori CR3 (CD11b/CD18), CR4 (CD11c/CD18) e il recettore per il C1q hanno un ruolo nel processo della fagocitosi.
- Il C3a, il C5a e il C4a hanno un’attività di anafilotossina. Essi provocano la degranulazione dei mastociti con conseguente aumento della permeabilità vascolare e contrazione della muscolatura liscia.
- Il C3b rivestendo i microrganismi migliora la loro fagocitosi.
- Il C3d aumenta la produzione di anticorpi da parte delle cellule B.
- Il C5a è un fattore chemiotattico per i neutrofili.
Difetti e deficit del sistema del complemento
Fondamentalmente i difetti e i deficit includono:
- Deficit di C1, C2, C3, MBL, MASP-2: suscettibilità alle infezioni batteriche ricorrenti.
- Deficit delle componenti C5, C9, fattore B, fattore D: suscettibilità alle infezioni da Neisseria.
- Difetti in C1, C4 e C5: lupus eritematoso sistemico.
- Difetti in CR2: immunodeficienza variabile comune.
- Deficit di CR3: deficit di adesitività leucocitari.
Giosuè Ruggiano
Fonti
- Immunità Innata: Il Paola Salvatore « Microbiologia e Immunologia « Scienze Biotecnologiche « Federica e-Learning (unina.it)
- Immunologia; malattie allergiche – Manuali MSD Edizione Professionisti (msdmanuals.com)
- https://en.wikipedia.org
- Immagine 1: Jules Bordet
- Microsoft PowerPoint –
- Immagine 2: Molecole solubili dell’immunità innata
- Immagine iniziale: IMMUNOLOGIA COMPLEMENTO.HTML