Caratteristiche generali dei recettori di membrana
I recettori di membrana, detti anche recettori transmembrana, sono costituiti da proteine intrinseche che possiedono tre diversi domini. Il dominio extracellulare situato esternamente alla cellula, ha la funzione di legare il ligando. Il dominio transmembrana, di solito ad alfa elica, è costituiti da amminoacidi idrofobici. Infine c’è il dominio intracellulare costituito da amminoacidi idrofili, interagisce con l’interno della cellula o dell’organello, trasmettendo il segnale.
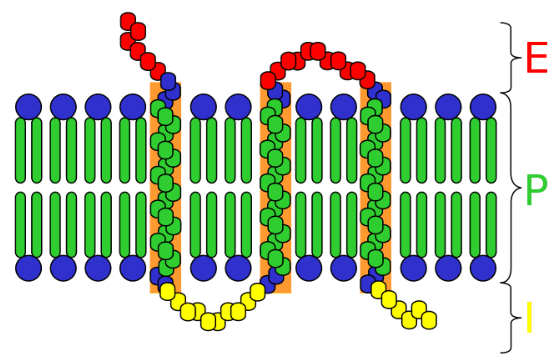
Entrano in gioco durante la comunicazione cellulare, rispondendo a determinati segnali che vengono captati proprio dai recettori di superficie.
Classificazione dei recettori di membrana
I recettori di membrana possono essere di due tipi:
- Ionotropici collegati a canali ionici
- Metabotropici: Si tratta di recettori che rispondono all’arrivo di ligandi extracellulari e danno il via ad una cascata di processi metabolici all’interno del citoplasma. Dal punto di vista funzionale possiamo classificarli in due grandi famiglie:
- Recettori accoppiati alle proteine G;
- Recettori accoppiati ad enzimi.
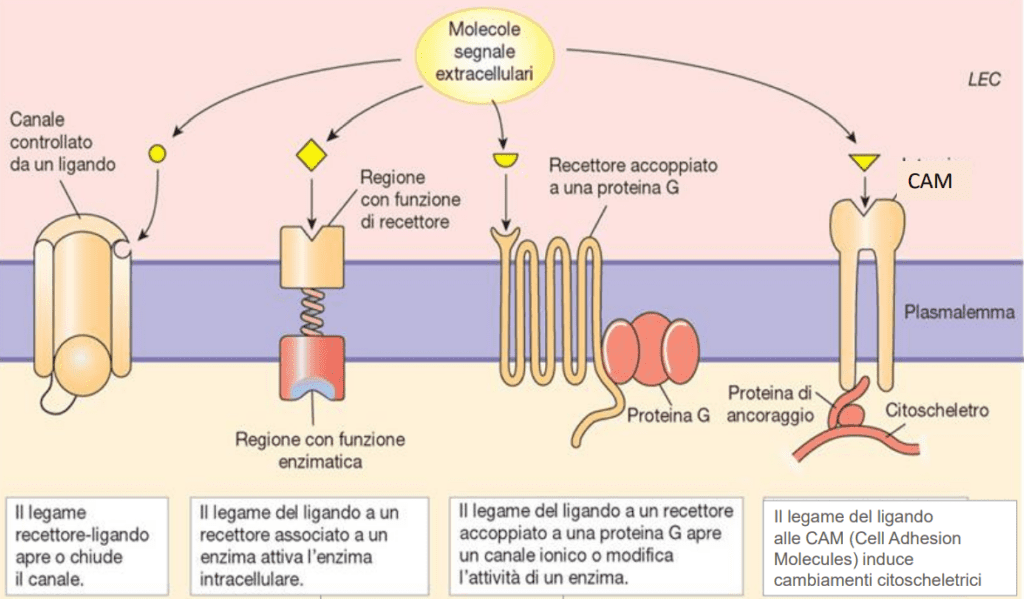
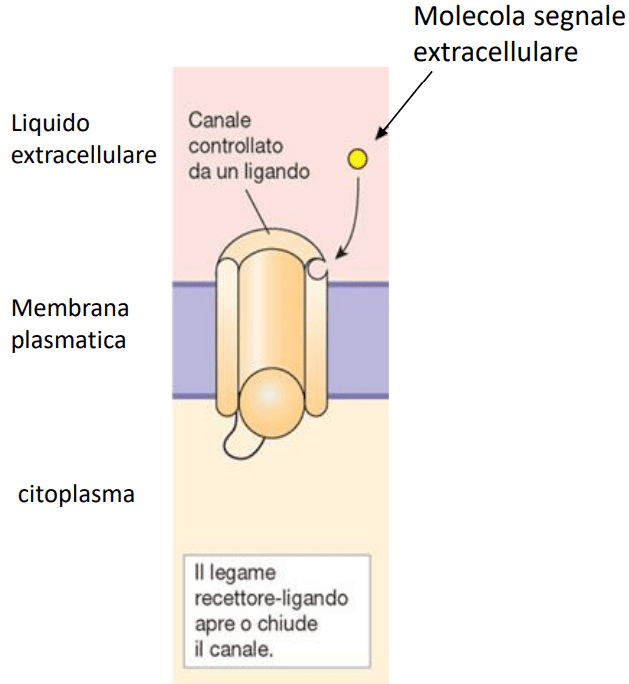
Recettori accoppiati a canali ionici
Questi recettori convertono i segnali chimici in segnali elettrici e sono presenti principalmente a livello di neuroni e cellule muscolari. La molecola di questi recettori presenta una porzione recettrice, esposta nel lato extracellulare e dotata dei siti di legame per la molecola del ligando. Inoltre possiedono una porzione effettrice, costituita dal canale ionico che attraversa lo spessore della membrana. Il complesso ligando recettore induce una variazione conformazionale del recettore con conseguente apertura del canale ionico a cui è associato, permettendo così il flusso degli ioni.
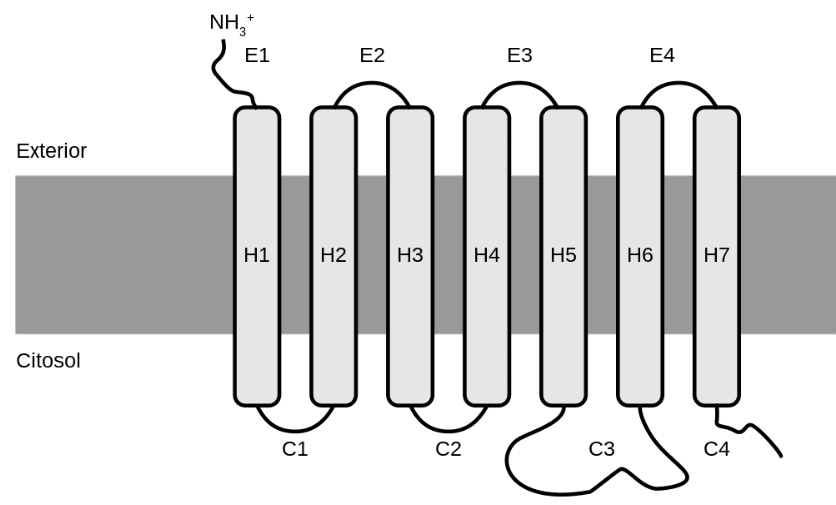
Recettori accoppiati a proteine G
I recettori accoppiati a proteine G (GPCR) sono proteine transmembrana costituite da 7 α-eliche connesse tra loro che, attraversando la membrana cellulare, interagiscono con il complesso delle proteine G. Le proteine G sono formate da 3 subunità che, allo stato inattivo, si presentano come subunità α e le subunità β e γ che formano un complesso tra loro. Nel momento in cui si forma il complesso ligando-recettore, il complesso βγ subisce una variazione conformazionale che lo porta ad associarsi ed attivare la proteina G. Ciò induce la dissociazione della subunità α dal complesso βγ e la riduzione dell’affinità della subunità α per il GDP, che viene sostituito da una molecola di GTP. A questo punto sia la subunità α che il complesso βγ possono interagire con le proteine bersaglio, che possono essere canali ionici o enzimi, che generano secondi messaggeri.
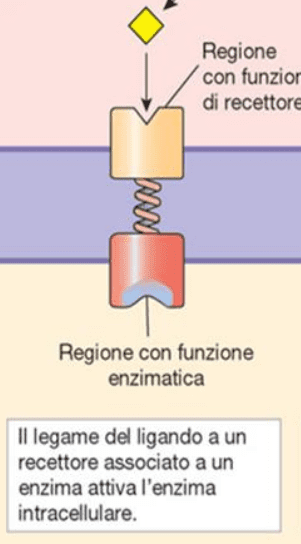
Recettori accoppiati ad enzimi
Questa classe di recettori sono proteine transmembrana che hanno un sito di legame esterno per la molecola segnale e un sito di legame interno per l’enzima. Possiedono due domini: un dominio di legame del ligando extracellulare e un dominio intracellulare, che ha una funzione catalitica. La molecola di segnalazione si lega al recettore all’esterno della cellula e provoca un cambiamento conformazionale sulla funzione catalitica situata sul recettore all’interno della cellula.
Trasporto attraverso la membrana
Le membrane cellulari hanno la funzione di creare barriere e rappresentano il “sistema” attraverso il quale la cellula crea dei compartimenti tra se stessa e l’ambiente esterno oppure tra un organello intracellulare e il citoplasma. Il doppio strato fosfolipidico, avendo una struttura fluida, è in grado di mediare alcuni tipi di trasporto cellulare. Esso inoltre forma compartimenti chiusi che si autoriparano, sigillandosi, in modo da mantenere intatta la propria integrità nel caso in cui intervengano lesioni di lieve o media entità.
Il trasporto di membrana può essere suddiviso in passivo o attivo in base alla richiesta di energia.
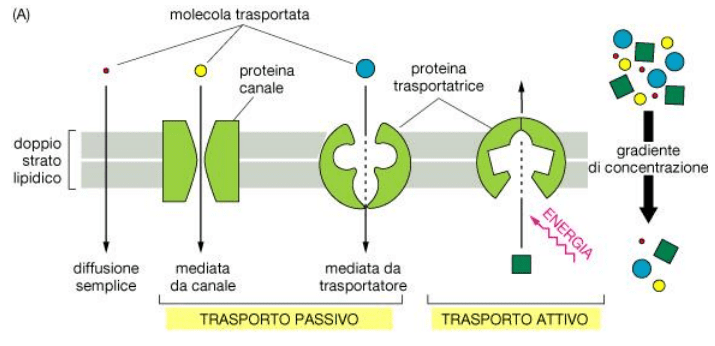
Trasporto passivo
Il trasporto passivo avviene secondo gradiente di concentrazione o elettrochimico, da una zona a maggiore concentrazione a zona a minore concentrazione, senza quindi richiesta di energia. Questo trasporto può avvenire per:
- Diffusione semplice;
- Diffusione facilitata;
- Diffusione attraverso i canali ionici.
Diffusione semplice
La diffusione semplice permette il passaggio di piccole molecole apolari (es. ossigeno) e molecole polari non cariche (es. acqua ed urea). Il flusso è secondo gradiente i concentrazione, e andrà da una zona più ricca di molecole ad una zona più povera.
Diffusione facilitata
Con la diffusione facilitata si ha il passaggio di sostanze nutritive all’interno della cellula, mediato da proteine transmembrana chiamate permeasi. Il passaggio avviene sempre secondo gradiente di concentrazione. Le tre caratteristiche degli scambi transmembrana mediati da proteine trasportatrici sono la specificità (ogni trasportatore opera solo su specifiche sostanze), la saturazione e la competizione (due sostanze affini competono per la proteina e l’una tende a deprimere il trasporto dell’altra).
Diffusione attraverso canali ionici
Il trasporto tramite canali ionici sono principalmente canali voltaggio-dipendenti (variazioni potenziale di membrana) e canali chemio-dipendenti. Presentano due caratteristiche fondamentali quali selettività (permeabilità solo a specifici ioni) e controllabilità (apertura o chiusura in base a specifici comandi).
Trasporto attivo
Il trasporto attivo è mediato da proteine transmembrana e avviene contro gradiente di concentrazione, per cui necessita di energia fornita da ATP. Le modalità di trasporto attivo attraverso membrana sono di tre tipi. La modalità uniporto consente il passaggio di una sola sostanza in un’unica direzione. Il simporto consente lo spostamento contemporaneo di due sostanze nella stessa direzione. Infine l’antiporto consente il passaggio di due sostanze in due direzioni opposte.
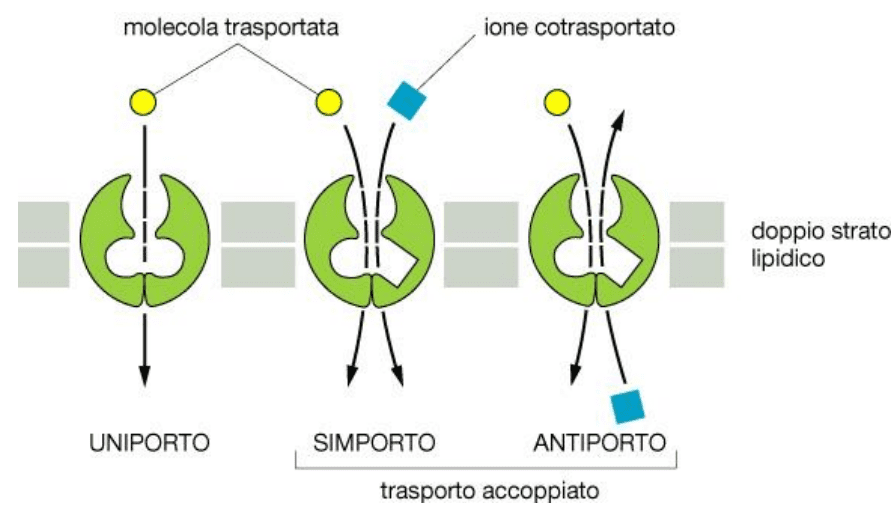
Il trasporto attivo può essere primario e secondario. Il trasporto attivo primario è mediato da proteine di membrana trasportatrici di soluti. Trasferisce le molecole contro gradiente di concentrazione, richiedendo quindi l’utilizzo di ATP. Nel trasporto secondario si ha il trasporto di una molecola contro gradiente che avviene grazie al precedente passaggio di un’altra molecola. Quindi si avvale inizialmente del trasporto attivo primario.
Fonti
- Fisiologia D’angelo e Peres Edi-ermes
- https://www.biopills.net
Crediti immagini
- Immagine in evidenza: https://www.biopills.net
- Figura 1: https://commons.wikimedia.org
- Figura 2: https://scienzemotorie.altervista.org
- Figura 3: https://www.unisalento.it
- Figura 4: https://commons.wikimedia.org
- Figura 5: https://www.unisalento.it
- Figura 6: http://www.med.unipg.it
- Figura 7: http://www.med.unipg.it